I AM PLEASED TO have an opportunity to take part in a lively and broad-ranging symposium dealing with the central theoretical principle of biology. I come here with much interest and anticipation, for if I could establish that the theory of evolution is invalid, it would be the greatest scientific discovery of the twentieth century. I would go down in history as one of the greatest savants of all time. A new era, a new age of science would be ushered in. Inconceivable wealth and power would be mine. If I could be even peripherally associated with such a monumental event, it would assure my future.
Theodosius Dobzhansky said in 1973, "Nothing in biology makes sense except in the light of evolution."{1} That assessment, coming from one of the leading biologists of the twentieth century, asserts that all of biological science collapses into a jumble of unrelated measurements and observations if not linked together by the central guiding generalization of evolution.
But biological science, chemistry, physics, and other branches of modern scientific inquiry are also joined together by a common purpose, rationale, and experimental approach. Thus a rejection of evolutionary theory would pull the rug out from under the whole edifice of modern science, allowing it to crash down in rubble all around us.
Because of all that hangs in the balance, it behooves us carefully to evaluate and question any attack on the theory of evolution, and to subject it to the most thorough scrutiny.
Professor Phillip Johnson's frontal assault on evolutionary theory, because of the substantial attention that it has received, deserves a careful critique, and if found unsatisfactory, deserves a measured rebuttal.
There is another reason, however, for a rigorous evaluation of evolutionary theory at this time Molecular biology has made tremendous strides in the last ten years, and these techniques and approaches have been applied to the design of evolutionary trees, analysis of relationships between taxonomic groups, measurements of chronological separation of species, comparison of genomes in diffenent taxonomic groups, and to a variety of issues in evolution. Workers in the field argue that this information imparts support and a new understanding of evolutionary theory, whereas Professor Johnson asserts that it merely shows similarities and has no bearing on the validity of evolutionary theory. In light of such conflicting views it is even more appropriate to reexamine the underpinnings of evolutionary theory at this time.
I do not wish to participate in a Johnson bashing, or a microanalysis of the arguments presented in his book. I do not believe that arguments over technical points in paleontology. molecular biology, or ecology will resolve the basic issue. Rather I wish to consider on a fundamental level the paramount philosophical question: the concept of purpose in biology.
The idea that the universe is ruled by a purposeful, guiding hand has been with us for a long time. Clearly this represents the primary issue with which we are concerned. Either mechanistic explanations are sufficient to explain the diversity, the function, and the beauty of the living world around us, or they are not.
But it is difficult to focus on the issue of purpose as it relates to a concept as broad and diverse as the study of evolutionary biology. For this reason I choose to examine the role of teleology in a much more restricted segment of biological science. which I feel qualified to discuss in detail and which is thoroughly understood on a molecular level, the field of immunology. I wish to focus particularly on the historical development of our understanding of how antibody molecules are generated. In doing so, l believe we can judge the current arguments concerning the adequacy of mechanistic evolutionary theory to explain the diversity of living forms.
Immunology, like other areas of biology, has developed rapidly in the last twenty years and has profited immensely from research using the techniques of molecular biology. In doing so, it has changed from a discipline that was mainly descriptive and that lacked clear models to a stage in which precise cause and effect relationships of the most minute events occurring within the immune system can be understood.
In the 1950s none of this was possible. It was known (and had been known for many years) that when humans or animals are exposed to foreign substances such as large protein or carbohydrate molecules (Usually through introduction into the circulation) they will produce a protective response in the form of circulating proteins. The foreign substance is known as an antigen, and the protein produced by the organism in response to it is known as an antibody. Antibodies are large molecules that constitute one of the main lines of defense against marauders from outside. Any question of their value for survival is answered by the pathology of AIDS in which the immune system is despoiled by HIV.
Moreover, these antibodies were known to combine with their respective inciting antigens in a reaction of great specificity. A lock and key mechanism was proposed by which the antibody molecule came precisely to fit the shape of the antigen. Following this binding it could, with the aid of other reactions, eliminate the offending antigen from the host.
Antibodies possessed an almost mystical quality. They were available in practically endless variety. For instance, Landsteiner had shown early in the century that an immune serum could distinguish between two proteins having as little as a single amino acid difference between them. Even finer levels of recognition were possible between D and L amino acids and ortho and para positions on benzene rings. This meant that the host must have the capacity to produce a virtually unlimited repertoire of responses. If one considers all the possible ways in which simple molecules can be modified and that the host should be able to generate antibodies to each of these modifications, then clearly there must be literally millions. and perhaps billions, of possible antibody types.
Three major theories were developed to explain how the tremendous diversity of the immune system could be generated. These three hypotheses were the instructive (or template) theory, the subcellular selection (or somatic mutation) theory, and the germ line theory. Elements of the latter two theories were later combined by Burnet in his "Clonal Selection Theory."
The instructive or template theory was proposed in its modern form in about 1930 by Felix Haurowitz and Linus Pauling (Mazunder, 1989; Kindt and Capra, 1984). it proposed that the antibodies were molecules that behaved in a plastic, flexible manner, so as to mold themselves to the shape of the antigen. Pauling, in 1940, suggested that the final step of synthesis of the antibody would be to fit the antigen. Thus the antibody was envisioned as an all-purpose, amorphous blob that after embracing the antigen was frozen into the mirror image of the antigen.
The template hypothesis was devised to get around the problem of storage of vast quantities of information. It got into trouble immediately, because it did not explain one of the most significant phenomena of immunology, that of vaccination. It had been recognized for hundreds of years that when an individual is exposed to a pathogenic agent such as smallpox virus, after a bout of disease the individual is immune to subsequent attacks of the same disease. Lady Mary Montagu, wife of the British ambassador to Constantinople in the early 1700s, is usually credited with bringing the discovery to England. She did this against the recommendation of her clergyman, who felt that vaccination against smallpox would be effective only in the heathen. In the nineteenth century Pasteur and others put the principle on a firm footing.
Thus if the instructional hypothesis were correct, it had to explain the fact that the immune system possessed a memory that could last sometimes for decades.
Its failure to explain immune memory satisfactorily cast the template theory in doubt from its inception. It was dealt a fatal blow by the new findings in the held of protein chemistry, which established that the folding of a protein molecule was a consequence of its amino acid content, which in turn was specified by its genetic program. Since the antibody molecule was composed of amino acids coded by its particular genetic message, there was no way in which it could change its shape, which was permanently fixed at its time of synthesis.
The second hypothesis was the somatic mutation hypothesis put forth in its modern form by Joshua Lederberg (1959) and others. This term refers to genetic modifications occurring in somatic or body cells, and not passed on through the germ line, i.e., the union of egg and sperm that binds each generation to the next. This hypothesis also possessed major drawbacks. It assumed that only a limited number of genes specified the structure of antibody molecules. These genes, however; were highly unstable and could mutate through an incredible variety of types, generating all possible antibody conformations. Thus in each generation the host would generate an entire panoply of antibodies, and the antigen would select and cause the amplification of the cell carrying its complementary antibody.
Although this theory escaped the problem of having to propose an antibody that could wrap around every conceivable antigen, it required two ad hoc assumptions: first, that antibody-forming cells were capable of exceedingly high mutation rates, and second, that the antigen could somehow select one cell, programmed to form the appropriate antibody from a great mass of cells producing irrelevant antibodies. Presumably following this selection there would be a stimulation of the relevant cell to generate a large population of descendants. At the time both of these problems were substantial roadblocks to an acceptance of the theory.
The third theory, known as the germ line theory (Kindt and Capra, 1984), proposed that all the information for producing all antibody types was carried in the genes, and passed from generation to generation through the germ line. Under the most direct form of this hypothesis, there would exist a separate gene for the antibody that reacts against every single molecule in the known universe. As I have suggested above, this would require an immense amount of genetic information. Some workers suggested at the time that this could account for the fact that mammals have so much more DNA than bacteria; i.e., aside from the housekeeping functions that they share with the bacteria, almost all their resources are given over to coding for antibodies.
The germ line theory also had a number of problems, not the least of which was the immense amount of baggage that an organism would be saddled with, and which, in the vast majority of instances, would never be called forth. Further, the theory was imbued with a strong element of teleology. It implied that mice, rabbits, and humans "knew" in some molecular sense that synthetic compounds would be invented millions of years before those compounds existed. By pursuing this line of reasoning, one would be forced to conclude that, even now, organisms carry with them the genetic blueprint for molecules that have yet to be synthesized in the laboratory of some as-yet-unborn organic chemist.
Between 1957 and 1959 F. MacFarlane Burnet proposed the clonal selection theory which opened the door to a resolution of these perplexing issues and to a modern understanding of antibody production (Ada, 1989). The crux of Burnet's theory was that in the animal there exist clones of cells which carry on their surfaces different recognition molecules. These molecules behave in a lock and key fashion, and bind the appropriate antigen. The reaction of the antigen with the recognition sites then activates that cell from a much larger population of clonal precursors, in effect selecting it. The selection process, through some unknown mechanism, then propels the cell down a long cascade of division and antibody production.
Burnet's proposal met with opposition from the start, and its vindication would require the advent of molecular approaches to immunology. As is so frequently true in the history of science, its final acceptance required concessions from both the germ line and somatic mutation factions, who lined up on either side of the debate (Silverstein, 1985). For the purposes of this discussion I will not follow the tortuous route through the next years that brought about our modern understanding of the mechanism of antibody diversity. During this intervening period of almost four decades, hundreds of thousands of years of investigators' time was spent working out the tine details of the immune system, which I can present here only in broadest outline, Although many details in immunology remain to be resolved, the mechanism of antibody diversity is well understood.
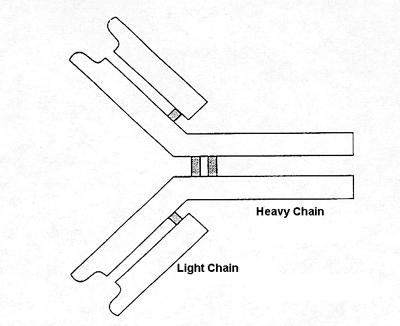
Antibody molecules share a common general structure, that of a "Y." with the portion that combines with the antigen being the ends of the prongs of the 'Y'' (Figure 1).
Figure 1. Four polypeptide chains, two light and two heavy, are joined together by disulphide bonds to form the antibody structure.
Each antibody molecule consists of two smaller units called light chains and two larger units known as heavy chains. The four submolecules are held together to form the complete, functional antibody molecule. Within the antibody-producing cell a number of genes are actually pulled out of their original position in the DNA and joined together to make the particular antibody molecule. There are four different families of genes. designated V, D, J, and C. A representative of each of these families will be extracted to form a composite gene in which a unique combination is assembled, like a hand at cards.
Antibody diversity arises at several levels. First, multiple variable genes are encoded in the germ lines, as had been predicted by the germ line theory, but there are only a few hundred at most to select from. Then a joining process takes place between the V, J, and D genes, which adds an additional level of variety. Next a recombinational inaccuracy can occur, such that the genes are not spliced together precisely, but can be joined at a variety of points. Then, as had been predicted by the somatic mutation hypothesis, the genes can undergo rapid fire mutations at a much higher frequency than the rest of the genome. Finally the heavy and light chains can get together in any combination, thus enormously amplifying the diversity.
So, following our Las Vegas analogy, antibody diversity is brought about by a molecular card deck that can be shuffled to generate a vast number of different antibody types. But once the players receive their hands, they can cheat and substitute one card for another through mutation. Thus each inveterate cellular gambler sits at the green felt molecular card table and gets a different hand from the deck, which he then further surreptitiously modifies in hopes of winning the jackpot of antibody production.
When invading proteins enter the body, they are recognized by the cells carrying antibodies reactive to them (Figure 2), and these cells are stimulated to proliferate and produce large quantities of antibody. The immune system functions in a fashion totally divorced from any guiding hand or purposeful control. It is a totally programmed response to environmental insult.
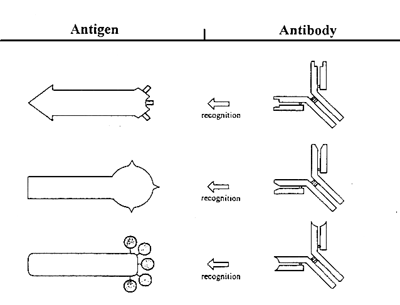
Figure 2. Antigens have characteristic regions, known as epitopes--molecular shapes that vary from antigen to antigen. Through recognition of antigen shapes, the generation of the appropriate antibodies is selected.
Are teleological explanations necessary in biology? We can see from this brief recounting of the development (or should I say evolution?) of a materialistic, mechanistic explanation for one of biology's cornerstones two important lessons for our consideration of the Darwinian debate; first. that selection determines the immune response, and second. that it is unnecessary to invoke teleological explanations to account for the phenomenon of antibody diversity.
In developing theories of the immune system, no less a biologist than the great Paul Ehrlich, founder of modern immunology, used the phrase "uralte protoplasma Weisheit,'' the "ancient wisdom of the protoplasm" (Silverstein, 1985). Ehrlich was proposing his side chain theory of antibody formation, and this ill-chosen term reflects, I believe, ambiguity that early biologists felt in designing mechanistic theories with which they were uncomfortable. Those early models were much too simple to account for the incredible complexity of biological systems. Ehrlich wrote the line in 1897, however, and in view of the vast gaps in his understanding, it is hardly surprising that teleologic phraseology crept into his descriptions.
At the time Ehrlich proposed his theory. it was thought that the only immunological response was to pathogenic organisms and toxic substances. These were believed to be few in number and thus the total number of different antibodies that an organism would be required to generate would be severely limited. It was unnecessary to propose a vast amount of unused antibody specificities. But as the immunological repertoire expanded to unmanageable proportions, it seemed that a Darwinian explanation for the existence of the immune system was no longer tenable. This ushered in the germ line theory with its immense and largely unused cornucopia of antibody types.
The guiding principle of Darwinian evolution convinced many investigators that the germ line theory, at least in its simplest form, had to be incorrect. It would make no sense for an organism to carry a vast amount of information that would never be employed. Surely it would be eliminated by natural selection. The development of our understanding of antibody diversity is a perfect example of the predictive power of the theory of evolution. If investigators had accepted a guiding, purposeful hand in the molding of the immune system, then the vast immune repertoire proposed by the germ line theory makes sense. It requires no explanation. The germ line theory is perfectly satisfactory, since the creator could look into the future and know what antibodies would be required.
But attention to the principle of natural selection forced a rejection of the germ line theory and a search for new experimental data. The present-day synthesis incorporating our knowledge of gene splicing, mutation, and rearrangement of the antibody-forming genes provides us with a completely rational and suitable account.
Thus in the development of immunological theories, the idea of purposeful design is no explanation at all, and is simply an excuse for muddled thinking.
If there is no scientific reason to evoke purpose in biology, does it have any place at all in science? Freeman Dyson (1979), a professor of physics at Princeton, has discussed the philosophical implications of his work in Disturbing the Universe. He has developed an interesting argument for a sort of purposeful spirit in the workings of the universe at the subatomic level. I think that this is a comforting notion, and is wholly consistent with the facts. Dyson, however, along with the vast majority of scientists, sees no reason for introducing the idea of purpose into biology.
The last hundred years and the work of thousands of investigators have established over and over again that mechanistic explanations are entirely adequate to explain the existence of the living world.
Descriptions based on mysticism, divine intervention, or purposeful guidance are untestable and provide no basis for an understanding of biological mechanisms. Purpose is a hindrance when introduced into discussions of biological phenomena, and can actually confuse us and impede our search for the truth.
Dyson, F. 1979. Disturbing theUniverse. Harper and Row, New York.
Futayma, D. 1982. Science on Trial. Pantheon Books, New York.
Kindt, T. J., and Capra, J. D. 1984. The Antibody Enigma. Plenum Press, New York.
Lederberg, J. 1959. Genes and antibodies. Science 129:1649.
Mazunder, P. H. 1989. The template theory of antibody formation and the chemical synthesis of the twenties. In Immunology: 19301980 (Ed. Mazander). Wall and Thompson, Toronto. pp. 13-33.
Silverstein, A. M. 1985. History of immunology: A history of the theories of antibody formation. Cellular Immunology 91:263-283.